![]()
![]()
![]()
Use LEFT and RIGHT arrow keys to navigate between flashcards;
Use UP and DOWN arrow keys to flip the card;
H to show hint;
A reads text to speech;
71 Cards in this Set
- Front
- Back
|
2.7.1 Somatosensorische waarneming
|
-tast, -druk, -proprioceptie, -warmte, -koude, -vibratie, -jeuk en -pijn.
Tactiele gewaarwordingen van tast en druk bieden informatie over het contact tussen ons lichaam en objecten uit de omgeving.
|
|
|
Tactiele gewaarwordingen van tast en druk bieden informatie over het contact tussen ons lichaam en objecten uit de omgeving.
Naast een belangrijke hulp in de objectidentificatie speelt de tastzin een belangrijke rol in |
de ontwikkeling en bij sociale interacties.
Proprioceptieve gewaarwordingen die worden veroorzaakt door de bewegingen van onze spieren en gewrichten leren ons iets over de positie en de bewegingen van ons lichaam.
Thermische sensaties als warmte of koude en de waarneming van pijn behoeden ons voor potentieel gevaarlijke omstandigheden en weefselbeschadiging.
De menselijke hand bevat niet alleen duizenden somatosensorische receptoren, maar is ook in constructief en motorisch opzicht bijzonder goed uitgerust om door middel van fijne manipulaties de oppervlakte en omtrekken van een object te onderzoeken. In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer de vingers over het object worden bewogen. |
|
|
Proprioceptieve gewaarwordingen die worden veroorzaakt door de bewegingen van onze spieren en gewrichten leren ons iets over |
de positie en de bewegingen van ons lichaam.
Thermische sensaties als warmte of koude en de waarneming van pijn behoeden ons voor potentieel gevaarlijke omstandigheden en weefselbeschadiging.
De menselijke hand bevat niet alleen duizenden somatosensorische receptoren, maar is ook in constructief en motorisch opzicht bijzonder goed uitgerust om door middel van fijne manipulaties de oppervlakte en omtrekken van een object te onderzoeken. In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer de vingers over het object worden bewogen. |
|
|
Thermische sensaties als warmte of koude en de waarneming van pijn behoeden ons voor |
potentieel gevaarlijke omstandigheden en weefselbeschadiging.
De menselijke hand bevat niet alleen duizenden somatosensorische receptoren, maar is ook in constructief en motorisch opzicht bijzonder goed uitgerust om door middel van fijne manipulaties de oppervlakte en omtrekken van een object te onderzoeken. In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer de vingers over het object worden bewogen. |
|
|
Functies van Tactiele gewaarwordingen:
-Tactiele gewaarwordingen van tast en druk bieden informatie over ….
-Naast een belangrijke hulp in de objectidentificatie speelt de tastzin een belangrijke rol in ...
-Proprioceptieve gewaarwordingen die worden veroorzaakt door de bewegingen van onze spieren en gewrichten leren ons iets over ….
-Thermische sensaties als warmte of koude en de waarneming van pijn behoeden ons voor ...
|
-Tactiele gewaarwordingen van tast en druk bieden informatie over het contact tussen ons lichaam en objecten uit de omgeving.
-Naast een belangrijke hulp in de objectidentificatie speelt de tastzin een belangrijke rol in de ontwikkeling en bij sociale interacties.
-Proprioceptieve gewaarwordingen die worden veroorzaakt door de bewegingen van onze spieren en gewrichten leren ons iets over de positie en de bewegingen van ons lichaam.
-Thermische sensaties als warmte of koude en de waarneming van pijn behoeden ons voor potentieel gevaarlijke omstandigheden en weefselbeschadiging. |
|
|
Functies van Tactiele gewaarwordingen kort:
O S P T |
- informatie over het contact tussen ons lichaam en objecten uit de omgeving. (objectidentificatie)
-de ontwikkeling en bij sociale interacties.
-de positie en de bewegingen van ons lichaam. (Proprioceptieve gewaarwordingen)
-waarneming van pijn behoeden ons voor potentieel gevaarlijke omstandigheden en weefselbeschadiging. (Thermische sensaties) |
|
|
|
één groot receptoroppervlak.
Zoals de macula het optisch centrum van de retina is, zo kunnen we onze handen als het tactiele centrum van ons lichaam beschouwen.
De menselijke hand bevat niet alleen duizenden somatosensorische receptoren, maar is ook in constructief en motorisch opzicht bijzonder goed uitgerust om door middel van fijne manipulaties de oppervlakte en omtrekken van een object te onderzoeken. In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer de vingers over het object worden bewogen. |
|
|
De menselijke hand bevat niet alleen duizenden somatosensorische receptoren, maar is ook in constructief en motorisch opzicht bijzonder goed uitgerust om door middel van fijne manipulaties |
de oppervlakte en omtrekken van een object te onderzoeken.
In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer de vingers over het object worden bewogen. |
|
|
In tegenstelling met de foveale perceptie die optimaal is wanneer het oog vrijwel onbeweeglijk blijft, is de tactiele perceptie het scherpst wanneer |
de vingers over het object worden bewogen. |
|
|
De informatie van de somatosensorische neuronen wordt via het ruggenmerg langs verschillende baansystemen naar de hersenen gevoerd.
De twee belangrijkste baansystemen zijn
D A
|
-het dorsolaterale kolom systeem, verantwoordelijk voor de transmissie van tast, druk en proprioceptie,
-en het fylogenetisch oudere anterolaterale systeem, verantwoordelijk voor de transmissie van pijn en temperatuursensaties. |
|
|
-het dorsolaterale kolom systeem is verantwoordelijk voor |
de transmissie van tast, druk en proprioceptie,
|
|
|
-het fylogenetisch oudere anterolaterale systeem is verantwoordelijk voor |
de transmissie van pijn en temperatuursensaties. |
|
|
Onderdelen:
t&d p of k p of n t
|
2.7.3 Proprioceptie of kinesthesie
|
|
|
2.7.2 Tast en druk
|
drukveranderingen van de huid.
Wanneer we in een hard object knijpen , zal de huid op onze vingertoppen een grotere weerstand ondervinden en onmiddellijk worden samengedrukt. Elastische objecten geven meer mee en zullen onze huid minder vervormen. Rekening houdend met de uitgeoefende kracht levert de mate waarin de huid wordt samengedrukt dus informatie over de stijfheid van het voorwerp. Wanneer we de vorm van een klein object willen voelen, kunnen we het object tussen duim en wijsvinger houden en het over onze vingertoppen laten rollen. Zachte, ronde objecten zullen teeds ongeveer dezelfde druk uitoefenen .
Een object met scherpe of onregelmatige randen zal daaren tegen abrupte drukveranderingen in de huid van de vingertoppen veroorzaken. De bewegende vingertoppen vertalen de omtrek van het voorwerp ineen specifiek patroon van huidvervormingen. Receptoren die in staat zijn om deze drukveranderingen te registreren, bieden belangrijke informatie over de vorm en de identiteit van het object.
|
|
|
2.7.3 Proprioceptie of kinesthesie
|
waar een lichaamsdeel zich bevindt zelfs wanneer bet een tijdlang niet werd bewogen. De proprioceptie is exact en precies, zij draagt bij tot vaardige en gerichte bewegingen.
|
|
|
Men zegt vaak dat pijn een lichaarnsbeschermende functie heeft. Pijn zorgt er immers voor dat |
we de hand tijdig van de hete kachel wegtrekken, dat we een opgelopen wonde onmiddellijk weten te lokaliseren en ze kunnen (laten) verzorgen.
Maar pijn heeft ook een andere zijde. Sommige levensbedreigende aandoeningen veroorzaken helemaal geen pijn en in andere gevallen kan een onuitstaanbare pijn geen enkele zinvolle functie meer vervullen (bijvoorbeeld fantoompijn). Voor mensen met chronische intense pijn is het pijngevoel vaak afschrikwekkender dan de dood zelf.
Dat pijn geen onfeilbaar systeem is, wordt door sommigen verklaard door het feit dat pijn teruggaat op een fylogenetisch zeer oude en nogal primitieve neuronale structuur (Beatty, 1995). Pijn is geen eenduidige reactie op weefselbeschadiging, maar is het product van een sensorische stimulus in interactie met andere, cerebrale invloeden die de beleving en de betekenis van pijnlijke sensaties mediëren.
Eenzelfde stimulus kan onder verschillende omstandigheden een spectrum van subjectieve re ponsen opleveren. Ook is pijn individueel bepaald (kleinze righeid) en blijken er zelfs culturele verschillen in de pijnbeleving te bestaan .
|
|
|
|
de temperatuur. Thermische receptoren liggen in kleine groepjes over het hele lichaam verspreid .
Het zijn slechts deze kleine vlekjes van 1 mm diameter die temperatuurveranderingen kunnen waarnemen. De vlekjes verschillen in reactie op bepaalde temperatuur schommelingen.
-De koudevlekjes reageren op een temperatuurdaling en resulteren in een gevoel van koude. -De warmtevlekjes reageren op een temperatuurverhoging en dragen bij tot een gevoel van warmte.
Wanneer een groter deel van het lichaam wordt blootgesteld aan een fysiologisch extreme temperatuur (bijvoorbeeld +45°C) is de overheersende gewaarwording die van een pijnlijke hitte. Wanneer echter een kleine hete stimulus selectief op een koudevlekje wordt aangebracht, is er een subjectieve sensatie van koude.
Dit paradoxaal effect wordt blijkbaar veroorzaakt doordat de koudereceptor wordt geprikkeld buiten zijn normaal fysiologisch temperatuurbereik . Een verhoogde respons van een koudereceptor wordt door de hersenen altijd geïnterpreteerd als koude.
Bij stimulatie van een grotere lichaamsoppervlakte worden uiteraard zowel warmte- als koudereceptoren gestimuleerd , wat in het brein tot een meer accurate waarneming van de huidtemperatuur leidt. |
|
|
Rekening houdend met de uitgeoefende kracht levert de mate waarin de huid wordt samengedrukt dus
|
informatie over de stijfueid van het voorwerp.
Wanneer we de vorm van een klein object willen voelen, kunnen we het object tussen duim en wijsvinger houden en het over onze vingertoppen laten rollen. Zachte, ronde objecten zullen teeds ongeveer dezelfde druk uitoefenen .
Een object met scherpe of onregelmatige randen zal daaren tegen abrupte drukveranderingen in de huid van de vingertoppen veroorzaken.
|
|
|
De bewegende vingertoppen vertalen de omtrek van het voorwerp ineen specifiek patroon van huidvervormingen.
Receptoren die in staat zijn om deze drukveranderingen te registreren, bieden belangrijke informatie over |
de vorm en de identiteit van het object.
|
|
|
De mechanische receptoren bestaan uit
d c a |
-een drukgevoelig uiteinde, -een cellichaam en -een axon.
Wanneer er druk op het sensibele uiteinde wordt uitgeoefend, reageert de cel door depolarisatie. Wanneer deze depolarisatie een bepaalde grens overschrijdt, ontstaat er een actiepotentiaal.
Een hogere druk resulteert in een sterkere depolarisatie en een snellere productie van actiepotentialen in het axon. De intensiteit van de stimulus kan ook worden afgeleid uit het aantal mechanische receptoren dat op de prikkel reageert.
Sterkere stimuli zullen ook de grenswaarden van minder gevoelige mechanische receptoren overschrijden. Uitgaande van de relatieve gevoeligheid van de reagerende receptoren kan ons zenuwstelsel de sterkte van de mechan ische tim ul us inschatten. Er bestaat een opvallende en lineaire gelijkenis tussen de intensiteit van de mechanische huidprikkels , de neuronale activiteit in de somatosensorische vezel baan en de bewuste perceptie van de stimulussterkte (Figu ur 2.26.). |
|
|
Wanneer er druk op het sensibele uiteinde wordt uitgeoefend, reageert de cel door |
depolarisatie. Wanneer deze depolarisatie een bepaalde grens overschrijdt, ontstaat er een actiepotentiaal.
Een hogere druk resulteert in een sterkere depolarisatie en een snellere productie van actiepotentialen in het axon. De intensiteit van de stimulus kan ook worden afgeleid uit het aantal mechanische receptoren dat op de prikkel reageert.
|
|
|
Een hogere druk resulteert in een sterkere depolarisatie en een snellere productie van actiepotentialen in het axon. De intensiteit van de stimulus kan ook worden afgeleid uit |
het aantal mechanische receptoren dat op de prikkel reageert.
Sterkere stimuli zullen ook de grenswaarden van minder gevoelige mechanische receptoren overschrijden. Uitgaande van de relatieve gevoeligheid van de reagerende receptoren kan ons zenuwstelsel de sterkte van de mechan ische tim ul us inschatten. Er bestaat een opvallende en lineaire gelijkenis tussen de intensiteit van de mechanische huidprikkels , de neuronale activiteit in de somatosensorische vezel baan en de bewuste perceptie van de stimulussterkte (Figu ur 2.26.). |
|
|
Sterkere stimuli zullen ook de grenswaarden van minder gevoelige mechanische receptoren overschrijden.
Uitgaande van de relatieve gevoeligheid van de reagerende receptoren kan ons zenuwstelsel de sterkte van de mechanische stimulus inschatten.
Er bestaat een opvallende en lineaire gelijkenis tussen |
de intensiteit van de mechanische huidprikkels , de neuronale activiteit in de somatosensorische vezel baan en de bewuste perceptie van de stimulussterkte (Figuur 2.26.). |
|
|
Er bestaat een opvallende en lineaire gelijkenis tussen de intensiteit van de mechanische huidprikkels , de neuronale activiteit in de somatosensorische vezel baan en de bewuste perceptie van de stimulussterkte (Figuur 2.26.). |
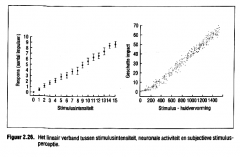
|
|
|
Mechanische receptoren verschillen van elkaar in de grootte van hun receptieve veld, hun gevoeligheid en hun mate van adaptatie.
Deze verschillende kwaliteiten kunnen uit de axonen van deze cellen worden afgeleid.
Adaptatie verwijst naar de mate waarin de cel reageert op temporele veranderingen van de stimulus.
-Traag adapterende vezels vertonen een…. -Snel adapterende vezels reageren slechts bij ...
|
-Traag adapterende vezels vertonen een verhoogde activatie zolang de drukstimulu aanwezig is. -Snel adapterende vezels reageren slechts bij het aanbrengen en het opnieuw verwijderen van de drukstimulus.
Zowel de trage als de snel adapterende vezels verschillen in de omtrek van hun receptieve velden. Puntcellen hebben smalle ovaalvormige receptieve velden met sterk gedefinieerde grenzen. Diffuse vezels hebben grote, onregelmatige en vage receptieve velden.
Verschillende types van mechanische receptorvezels zijn derhalve in staat om verschillende aspecten van somatische stimulatie te signaleren. |
|
|
Zowel de trage als de snel adapterende vezels verschillen in de omtrek van hun receptieve velden. Puntcellen hebben smalle ovaalvormige receptieve velden met sterk gedefinieerde grenzen. Diffuse vezels hebben grote, onregelmatige en vage receptieve velden.
Verschillende types van mechanische receptorvezels zijn derhalve in staat om |
verschillende aspecten van somatische stimulatie te signaleren. |
|
|
Er zijn twee grote klassen van mechanische receptoren :
|
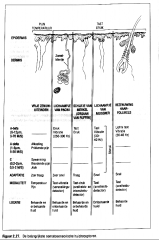
-de vrije en -de ingekapselde zenuwuiteinden
(Figuur 2.27.).
|
|
|
-de vrije en -de ingekapselde zenuwuiteinden |
-Bij de vrije zenuwuiteinden is de drukgevoelige zone niet omgeven door een of andere cellulaire structuur. De vrije zenuwuiteinden bezenuwen de huid en aanverwante structuren (bijvoorbeeld de haartjes) en zijn vrijwel over het gehele lichaam terug te vinden (tot in de cornea van het oog).
-De ingekapselde zenuwuiteinden zijn omgeven door meer gecompliceerde mechanische structuren van bindweefsel.
De lichaampjes van Pacini zijn de meest bekende ingekapselde zenuwuiteinden. |
|
|
De lichaampjes van Pacini zijn de meest bekende ingekapselde zenuwuiteinden. Ze zijn vrij groot (1 tot 4 mm) en bestaan uit een concentrische reeks van met vloeistof gevulde capsules die het centraal gelegen zenuwuiteinde omsluiten.
Wat gebeurt er als er druk uitgeoefend wordt op de lichaampjes van Pacini?
|
-Wanneer druk op een lichaampje van Pacini wordt uitgeoefend, verplaatst de druk in de capsules zich zodanig dat de druk op het zenuwuiteinde vermindert.
-Wanneer de druk verdwijnt, stroomt de vloeistof terug naar zijn oorspronkelijke plaats , wat de cel opnieuw exciteert.
-Wanneer de druk constant blijft, wordt het zenuwuiteinde niet geprikkeld.
De lichaampjes van Pacini reageren dus enkel op drukveranderingen, niet op de constante aan- of afwezigheid van druk; het zijn dus hoog adaptieve receptoren.
|
|
|
Wat zijn de lichaampjes van Pacini dus? |
De lichaampjes van Pacini reageren dus enkel op drukveranderingen, niet op de constante aan- of afwezigheid van druk;
het zijn dus hoog adaptieve receptoren. |
|
|
Naast de lichaampjes van Pacini onderscheiden we vooral
m r m |
-de lichaampjes van Meissner, -de eindorgaantjes van Ruffini en -de schijfjes van Merkel.
Een lichaampje van Meissner wordt bezenuwd door tot wel zes gemyeliniseerde zenuwvezels. Ze bevinden zich vooral in de gevoelige, haarloze huid van de vingers. Ze reageren bij voorkeur op een vluchtige stimulatie zoals wanneer men met de vingertoppen over een voorwerp wrijft. De lichaampjes van Pacini en Meissner worden vooral door de snel adapterende vezels bezenuwd.
De eindorgaantjes van Ruffini worden vooral in de behaarde huid aangetroffen en zijn net zoals de schijfjes van Merkel vooral actief bij een constante vervorming van de huid. Deze laatste twee receptortypes worden dan ook vooral door traag adapterende vezels bezenuwd. |
|
|
Een lichaampje van Meissner wordt bezenuwd door tot wel zes gemyeliniseerde zenuwvezels. Ze bevinden zich vooral in |
de gevoelige, haarloze huid van de vingers. Ze reageren bij voorkeur op een vluchtige stimulatie zoals wanneer men met de vingertoppen over een voorwerp wrijft. De lichaampjes van Pacini en Meissner worden vooral door de snel adapterende vezels bezenuwd.
|
|
|
De eindorgaantjes van Ruffini worden vooral in de behaarde huid aangetroffen en zijn net zoals de schijfjes van Merkel vooral actief bij |
een constante vervorming van de huid. Deze laatste twee receptortypes worden dan ook vooral door traag adapterende vezels bezenuwd. |
|
|
De belangrijkste somatosensorische huidcellen |
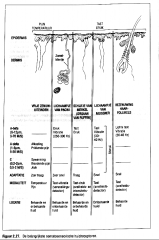
|
|
|
De axonen van de meeste mechanische receptoren maken contact met het centraal zenuwstelsel via het ruggenmerg (of de craniale zenuwen).
Het ruggenmerg is een structuur van |
stijgende en dalende vezel die de hele wervelkolom vult.
Toch bestaat er in het ruggenmerg een segmentale organisatie die wordt veroorzaakt doordat de afferente prikkels het ruggenmerg enkel kunnen binnendringen door 31 paar (linker en rechter) dorsale wortels via uitsparingen tussen de ruggenwervels .
|
|
|
Toch bestaat er in het ruggenmerg een segmentale organisatie die wordt veroorzaakt doordat de |
afferente prikkels het ruggenmerg enkel kunnen binnendringen door 31 paar (linker en rechter) dorsale wortels via uitsparingen tussen de ruggenwervels . |
|
|
Elke dorsale wortel bedient een welbepaalde regio van het lichaam : |
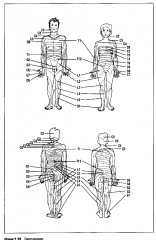
een dermatoom (Figuur 2.28.). |
|
|
Hoewel naburige dermatomen elkaar enigszins overlappen , bieden zij een duidelijke segmentale organisatie van het perifere gedeelte van het omatosensorisch systeem.
In elke dorsale wortel komen de verschillende vezeltypes samen die afkomstig zijn van hetzelfde dermatoom. Bij het binnendringen van het ruggenmerg splitsen de verschillende vezeltypes zich op in verschillende stijgende banen die |
afhankelijk van hun functionele rol afzonderlijke anatomische routes volgen
(zie 'De spinale zenuwen' in het eerste hoofdstuk). |
|
|
Sensorische prikkels van tast en druk reizen via het …. naar het brein. |
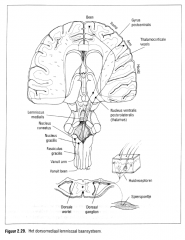
dorsomediaal lemniscaal baan systeem naar bet brein
('lemniscus' betekent bundel) (Figuur 2.29.).
|
|
|
De cellichamen van de perifere mechanische receptoren liggen dicht bij het ruggenmerg in groepjes bij elkaar, de …. |
dorsale ganglia.
Deze cellichamen hebben enkel een metabolische functie. De vezels van de cellichamen zijn lang en goed gemyeliniseerd en voorzien in een snelle transmissie van de actiepotentialen .
De vezels dringen de wervelkolom binnen via de dorsale wortels en vormen de stijgende dorsale kolom . Zij synapteren in de dorsale kolomkemen ter hoogte van de medulla oblongata. De dorsale kolom is een belangrijk baansysteem in het ruggenmerg. Inzijn stijgende projectie worden de dorsale wortelvezels van de opeenvol gende dermatomen lateraal aan dit baansysteem toegevoegd.
De dorsale kolom verbreedt zich dus in caudocraniale richting waarbij informatie uit caudale delen van het lichaam zich meer mediaal en informatie uit craniale delen zich meer lateraal bevindt. Op deze manier bewaart de dorsale kolom een somatotopische organisatie.
Deze somatotopische organisatie handhaaft zich in de dorsale kolornkernen. De mediale nucleus gracilis ontvangt de projecties van het onderste lichaamsgedeelte, de laterale nucleus cuneatus ontvangt de projecties van het bovenste lichaamsgedeelte.
De vezels uit beide dorsale kolomkernen kruisen de middellijn en projecteren samen als de lemniscus medialis naar de posterieure ventrolaterale kern van de thalamus. Vanuit deze tbalamische kern en de posterieure ventromediale tbalam uskem (die zijn input van de craniale trigeminus zenuw ontvangt die de lip en mondarea bezenu wt) wordt de somatosensorische informatie op de primaire somatosensorische cortex geprojecteerd. |
|
|
De dorsale ganglia.
Deze cellichamen hebben enkel een metabolische functie. De vezels van de cellichamen zijn lang en goed gemyeliniseerd en voorzien in een snelle transmissie van de actiepotentialen .
De vezels dringen de wervelkolom binnen via |
de dorsale wortels en vormen de stijgende dorsale kolom . Zij synapteren in de dorsale kolomkernen ter hoogte van de medulla oblongata.
De dorsale kolom is een belangrijk baansysteem in het ruggenmerg. In zijn stijgende projectie worden de dorsale wortelvezels van de opeenvolgende dermatomen lateraal aan dit baansysteem toegevoegd.
De dorsale kolom verbreedt zich dus in caudocraniale richting waarbij informatie uit caudale delen van het lichaam zich meer mediaal en informatie uit craniale delen zich meer lateraal bevindt. Op deze manier bewaart de dorsale kolom een somatotopische organisatie.
Deze somatotopische organisatie handhaaft zich in de dorsale kolornkernen. De mediale nucleus gracilis ontvangt de projecties van het onderste lichaamsgedeelte, de laterale nucleus cuneatus ontvangt de projecties van het bovenste lichaamsgedeelte.
De vezels uit beide dorsale kolomkernen kruisen de middellijn en projecteren samen als de lemniscus medialis naar de posterieure ventrolaterale kern van de thalamus. Vanuit deze tbalamische kern en de posterieure ventromediale tbalam uskem (die zijn input van de craniale trigeminus zenuw ontvangt die de lip en mondarea bezenu wt) wordt de somatosensorische informatie op de primaire somatosensorische cortex geprojecteerd. |
|
|
De dorsale kolom is een belangrijk baansysteem in het ruggenmerg. In zijn stijgende projectie worden de dorsale wortelvezels van de opeenvolgende dermatomen lateraal aan dit baansysteem toegevoegd.
De dorsale kolom verbreedt zich dus in |
caudocraniale richting waarbij informatie uit caudale delen van het lichaam zich meer mediaal en informatie uit craniale delen zich meer lateraal bevindt. Op deze manier bewaart de dorsale kolom een somatotopische organisatie.
Deze somatotopische organisatie handhaaft zich in de dorsale kolornkernen. De mediale nucleus gracilis ontvangt de projecties van het onderste lichaamsgedeelte, de laterale nucleus cuneatus ontvangt de projecties van het bovenste lichaamsgedeelte.
De vezels uit beide dorsale kolomkernen kruisen de middellijn en projecteren samen als de lemniscus medialis naar de posterieure ventrolaterale kern van de thalamus. Vanuit deze tbalamische kern en de posterieure ventromediale tbalam uskem (die zijn input van de craniale trigeminus zenuw ontvangt die de lip en mondarea bezenu wt) wordt de somatosensorische informatie op de primaire somatosensorische cortex geprojecteerd. |
|
|
Deze somatotopische organisatie handhaaft zich in de dorsale kolomkernen. De mediale nucleus gracilis ontvangt |
de projecties van het onderste lichaamsgedeelte, de laterale nucleus cuneatus ontvangt de projecties van het bovenste lichaamsgedeelte.
De vezels uit beide dorsale kolomkernen kruisen de middellijn en projecteren samen als de lemniscus medialis naar de posterieure ventrolaterale kern van de thalamus. Vanuit deze tbalamische kern en de posterieure ventromediale tbalam uskem (die zijn input van de craniale trigeminus zenuw ontvangt die de lip en mondarea bezenu wt) wordt de somatosensorische informatie op de primaire somatosensorische cortex geprojecteerd. |
|
|
De vezels uit beide dorsale kolomkernen kruisen de middellijn en projecteren samen als de lemniscus medialis naar de posterieure ventrolaterale kern van de thalamus.
Vanuit deze thalamische kern en de posterieure ventromediale thalamuskem (die zijn input van de craniale trigeminus zenuw ontvangt die de lip en mondarea bezenuwt) wordt de somatosensorische informatie op de …….geprojecteerd. |
de primaire somatosensorische cortex geprojecteerd. |
|
|
Aangezien het lemniscaal baansysteem de middellijn kruist, ontvangt de primaire somatosensorische area zijn informatie uit de contralaterale lichaamshelft.
De primaire somatosensorische area is gelegen in de postcentrale gyrus van de pariëtale lob, de Brodmann-areas 1, 2, 3a en 3b. Elk van deze areas heeft niet alleen een verschillende cytoarchitecturale structuur, maar blijkt ook functioneel gespecialiseerd.
|
-Area 3a ontvangt informatie van mechanische receptoren uit het spierweefsel. -Areas 1 en 3b ontvangen input van huidreceptoren. -In area 1 komt ook de informatie vanuit de lichaampjes van Pacini toe, wat niet het geval is in area 3b. -Mechanische receptoren gelegen in de gewrichten en diepere weefsels projecteren naar area 2. |
|
|
Binnen het gehele baansysteem blijft de somatotopische organisatie strikt gehandhaafd.
Maar ook hier stellen we, net zoals de middenfrequenties bij de auditus en het maculaire optische centrum bij de visus, wat vast?
|
een corticale uitvergroting van de meer belangrijke somatosensorische regio 's vast.
Gevoelige delen van ons lichaam (lippen, tong, vingers) krijgen een relatief groter aandeel van de cortex in vergelijking met de minder gevoelige delen van het lichaam (rug, romp).
|
|
|
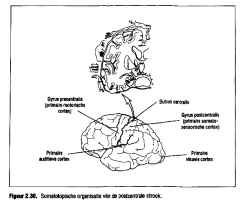
Figuur 2.30. toont de somatotopische organisatie van de postcentrale trook bij de mens.
Let op de relatief grote gebieden voor de gevoeligheid van de tong (spraak) en de oppositionele duim (objectmanipulatie). We vinden deze somatotopische organisatie terug in de vier smalle parallel lopende areas (1, 2, 3a en 3b) die elk verschillende somatosensorische informatie verwerken (Kaas, Nelson, Sur & Marzenich, 1981). |
|
|
|
Het receptief veld van een somatosensorisch neuron definiëren we als het gedeelte van de huidoppervlakte dat, indien gestimuleerd:
|
de vezel activeert.
De cellen van de postcentrale strook hebben sterk gedefinieerde receptieve velden. De grootte van deze velden is erg variabel en |
|
|
De cellen van de postcentrale strook hebben sterk gedefinieerde receptieve velden. De grootte van deze velden is erg variabel en afhankelijk van hun initiële locatie op het lichaam.
Lichaamsgebieden waar veel mechanische receptoren zitten en waar we dus een hoge gevoeligheid kennen (zoals tong en vingers), hebben |
zeer kleine receptieve velden.
Gebieden met weinig receptoren (rug, romp, benen) hebben grote receptieve velden (Werner & Whitsel, 1973).
Ook de neocorticale somatosensorische cortex blijkt georganiseerd in functionele kolommen (Mount castle, 1957). Wanneer een elektrode loodrecht in de primaire sensorische cortex wordt ingebracht, vertonen de boven en onder elkaar gelegen neuronen dezelfde eigenschappen, zoals een voorkeursreactie op lichte aanraking maar niet op diepe gevoeligheid of vibratie.
Penetratie van een ander cordiaal gebied toont neuronen die gevoelig zijn voor temperatuur, maar niet voor tast en druk. Een schuin ingebrachte elektrode die verschillende functionele kolommen kruist, vindt neuronen met een verschillend soort van voorkeurs stimulatie. |
|
|
Ook de neocorticale somatosensorische cortex blijkt georganiseerd in functionele kolommen (Mount castle, 1957).
|
-Wanneer een elektrode loodrecht in de primaire sensorische cortex wordt ingebracht, vertonen de boven en onder elkaar gelegen neuronen dezelfde eigenschappen, zoals een voorkeursreactie op lichte aanraking maar niet op diepe gevoeligheid of vibratie.
-Penetratie van een ander cordiaal gebied toont neuronen die gevoelig zijn voor temperatuur, maar niet voor tast en druk.
-Een schuin ingebrachte elektrode die verschillende functionele kolommen kruist, vindt neuronen met een verschillend soort van voorkeurs stimulatie. |
|
|
|
de gewaarwording van de eigen lichaamshouding en beweging.
|
|
|
Proprioceptieve gewaarwordingen vertonen weinig adaptatie, we weten altijd waar een lichaamsdeel zich bevindt zelfs wanneer bet een tijdlang niet werd bewogen.
De proprioceptie is exact en precies, zij draagt bij tot |
vaardige en gerichte bewegingen. |
|
|
Proprioceptie begint bij de |
somatosensorische receptoren in de gewrichten en skelet spieren.
|
|
|
Gewrichtsreceptoren zijn gespecialiseerde mechanische receptoren die zodanig in de gewrichten geplaatst zijn dat zij de hoek waarin het gewricht zich bevindt, kunnen detecteren.
We beschikken over vier types gewrichtsreceptoren die elk een verschillende anatomische structuur en functionele eigenschappen bezitten . |
Proprioceptieve informatie komt ook van receptoren die in de skeletspieren gelegen zijn. Spierspoeltjes zijn kleine receptoren die zowel door sensorische (afferente) als motorische (efferente) vezels worden bezenuwd.
Zij spelen een belangrijke rol in de regulatie van spierbewegingen, maar bieden tevens accurate informatie over de lengte (spanning versus ont panning) van de spier en dragen derhalve ook bij tot de proprioceptie . |
|
|
|
een motiverende (en emotionele) functie en daarin verschilt pijn wezenlijk van de andere zintuigen.
Men zegt vaak dat pijn een lichaarnsbeschermende functie heeft. Pijn zorgt er immers voor dat we de hand tijdig van de hete kachel wegtrekken, dat we een opgelopen wonde onmiddellijk weten te lokaliseren en ze kunnen (laten) verzorgen. |
|
|
Maar pijn heeft ook een andere zijde.
Sommige levensbedreigende aandoeningen veroorzaken helemaal geen pijn en in andere gevallen kan een onuitstaanbare pijn geen enkele zinvolle functie meer vervullen (bijvoorbeeld fantoompijn). Voor mensen met chronische intense pijn is het pijngevoel vaak afschrikwekkender dan de dood zelf.
Dat pijn geen onfeilbaar systeem is, wordt door sommigen verklaard door het feit dat |
pijn teruggaat op een fylogenetisch zeer oude en nogal primi tieve neuronale structuur (Beatty, 1995).
Pijn is geen eenduidige reactie op weefselbeschadiging, maar is het product van een sensorische stimulus in interactie met andere, cerebrale invloeden die de beleving en de betekenis van pijnlijke sensaties mediëren.
Eenzelfde stimulus kan onder verschillende omstandigheden een spectrum van subjectieve re ponsen opleveren. Ook is pijn individueel bepaald (kleinzierigheid) en blijken er zelfs culturele verschillen in de pijnbeleving te bestaan . |
|
|
Pijmeceptoren of nociceptoren (van het Latijnse nocere : verwonden) liggen over het hele lichaam verspreid. Nociceptoren zijn allemaal vrije zenuwuiteinden.
Ondanks hun anatomische gelijkenis vertonen ze een verschillende voorkeur voor bepaalde fysische stimuli . |
-Mechanische nociceptoren reageren voornamelijk op een intense mechanische stimu latie (scherpe objecten). -Warmtenociceptoren reageren vooral op hitte . -Andere nociceptoren reageren bij voorkeur op chemische substanties zoals kalium, dat vrijkomt bij celbeschadiging. |
|
|
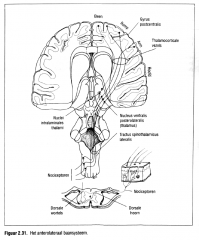
De pijnprikkels volgen een andere, fylogenetisch oudere, anterolaterale baan naar het centraal zenuwstelsel (Figuur 2.31.). |
|
|
|
Nociceptoren communiceren met het ruggenmerg door middel van twee soorten vezels met een verschillende geleidingssnelheid. |
-A-delta-vezels -ongemyelinieerde C-vezels
A-delta-vezels zijn bredere gemyeliniseerde vezels met een hoge geleidingssnelheid (5 tot 30 meter per seconde). Activatie van de A-deltavezels produceert een snelle, felle, goed lokaliseerbare pijn .
De dunnere, ongemyelinieerde C-vezels hebben een tragere geleidingssnelheid (0,5 tot 2 meter per seconde) en zijn verantwoordelijk voor de trage, doffe, minder gelokaliseerde pijn waarneming. |
|
|
Zowel de A-delta- als de C-vezels dringen het ruggenmerg binnen via de dorsale
|
-de ipsilaterale dorsale hoornen (het dorsale deel van de vlinder vormige grijze stof in het ruggenmerg) , -kruisen vrijwel integraal de middellijn (een kleine ipsilaterale projectie wordt vermoed) , -en stijgen via de laterale spinothalamische baan naar de thalamus, -via de spinoreticulaire baan naar de formatio reticularis in de hersen stam en -via de spinotectale baan naar de colliculi superior en inferior en de periaqueductale grijze stof , een neuroncomplex gelegen in het midden van de hersenstam.
Slechts een klein aantal vezels projecteert via de thalarnus naar de neocortex (Kelly, 1985). De cerebrale cortex zelf blijkt dus weinig betrokken bij de pijnwaarneming. |
|
|
Slechts een klein aantal vezels projecteert via de thalarnus naar de neocortex (Kelly, 1985). De cerebrale cortex zelf blijkt dus weinig betrokken bij ... |
de pijnwaarneming. |
|
|
|
de temperatuur.
Thermische receptoren liggen in kleine groepjes over het hele lichaam verspreid . Het zijn slechts deze kleine vlekjes van 1 mm diameter die temperatuurveran deringen kunnen waarnemen.
|
|
|
De vlekjes verschillen in reactie op bepaalde temperatuur schommelingen.
-De koudevlekjes reageren op ... -De warmtevlekjes reageren op ...
|
-De koudevlekjes reageren op een temperatuurdaling en resulteren in een gevoel van koude. -De warmtevlekjes reageren op een temperatuurverhoging en dragen bij tot een gevoel van warmte. |
|
|
Wanneer een groter deel van het lichaam wordt blootgesteld aan een fysiologisch extreme temperatuur (bijvoorbeeld +45°C) is de overheersende gewaarwording die van een pijnlijke hitte.
Wanneer echter een kleine hete stimulus selectief op een koudevlekje wordt aangebracht, is er een subjectieve sensatie van koude.
Dit paradoxaal effect wordt blijkbaar veroorzaakt doordat |
de koudereceptor wordt geprikkeld buiten zijn normaal fysiologisch temperatuurbereik . Een verhoogde respons van een koudereceptor wordt door de hersenen altijd geïnterpreteerd als koude.
Bij stimulatie van een grotere lichaamsoppervlakte worden uiteraard zowel warmte- als koudereceptoren gestimuleerd , wat in het brein tot een meer accurate waarneming van de huidtemperatuur leidt. |
|
|
Een verhoogde respons van een koudereceptor wordt door de hersenen altijd geïnterpreteerd als koude.
Bij stimulatie van een grotere lichaamsoppervlakte worden uiteraard zowel warmte- als koudereceptoren gestimuleerd , wat in het brein tot … leidt. |
een meer accurate waarneming van de huidtemperatuur leidt. |
|
|
De neuronen van de temperatuurgevoelige vrije zenuwuiteinden hebben ook A-delta of C-axonen.
Warmte wordt vrijwel geheel gemedieerd door
|
de ongemyeliniseerde C-vezels.
Deze cellen verhogen hun vuursnelheid met een stijgende temperatuur in het fysiologische 'warme' bereik.
|
|
|
Bij hogere temperaturen (boven 45°C) ervaren we de temperatuur als een pijnlijke hitte.
Bij deze temperatuur komt het systeem van de …….in het spel. |
warmtenociceptoren of gemyeliniseerde A-deltavezels in het spel.
De A-deltavezels vuren in verhouding tot de intensiteit van de pijnlijke warmtestimuli. |
|
|
Koudereceptoren gebruiken zowel |
A-delta- als C-vezels
en signaleren koude beneden een fysiologisch en psychologisch neutraal punt.
In vergelijking met de hoge resolutie van de waarneming van tast en druk is de spatiële, lokalisatorische capaciteit van de thermoreceptoren bijzonder gering.
Vermoedelijk zijn de receptieve velden van de temperatuurwaarneming zeer groot. |
|
|
In vergelijking met de hoge resolutie van de waarneming van tast en druk is de spatiële, lokalisatorische capaciteit van de thermoreceptoren |
bijzonder gering.
Vermoedelijk zijn de receptieve velden van de temperatuurwaarneming zeer groot.
Beide receptoren gebruiken A-delta- en C-vezels, en beide gebruiken ze het anterolateraal systeem in hun projectie naar de hersenstam en de thalamus. Voor temperatuur werd totnogtoe geen corticale representatie gevonden . |
|
|
Vermoedelijk zijn de receptieve velden van de temperatuurwaarneming zeer groot.
|
het pijnbaan systeem.
Beide receptoren gebruiken A-delta- en C-vezels, en beide gebruiken ze het anterolateraal systeem in hun projectie naar de hersenstam en de thalamus. Voor temperatuur werd totnogtoe geen corticale representatie gevonden . |
|
|
Het thermoreceptieve baan systeem en het pijnbaan systeem gebruiken beide... |
A-delta- en C-vezels,
en beide gebruiken ze het anterolateraal systeem in hun projectie naar de hersenstam en de thalamus.
Voor temperatuur werd totnogtoe geen corticale representatie gevonden . |